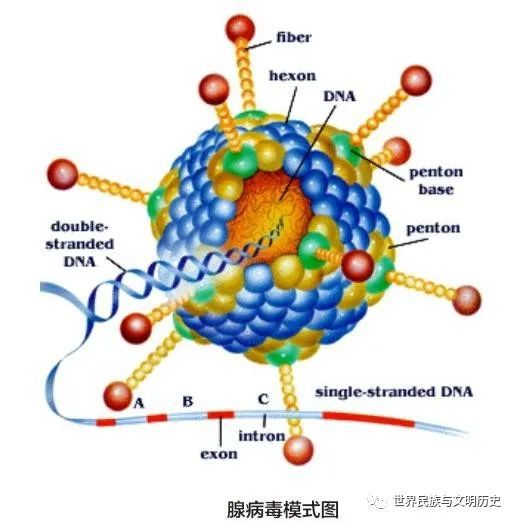
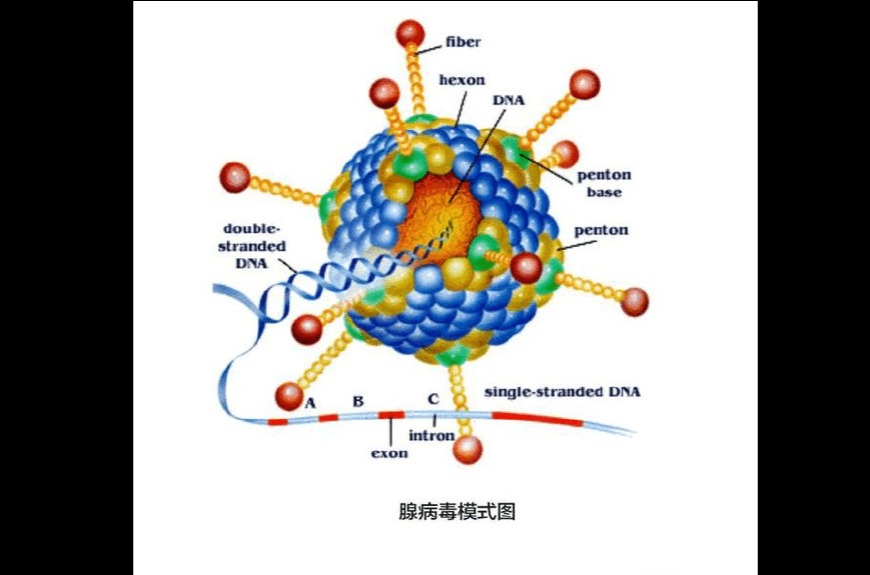
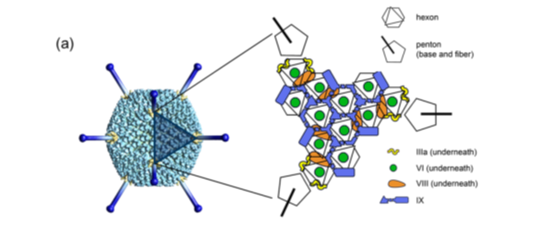
腺病毒是一种无外壳的双链DNA病毒,基因组长约36kb,衣壳()呈规则的20面体结构,直径约80-110nm。衣壳含有240个联体(hexon)、12个联体()及12根纤毛(fiber),除此此外也有其他一些小蛋白,如VI、VIII、IX、IIIa和IVa2等。六联体是产生病毒衣壳20个三角形面的主要蛋白,12个顶端是5个联体亚单位和3个纤毛蛋白组成的复合物,12根纤毛以五联体蛋白为基底由衣壳表面伸出腺病毒包装技术,纤毛顶部形成头节区(knob)。五联体和纤毛的头节区可与细胞表层的细菌受体结合,在真菌感染细胞过程中起着十分重要的功效。腺病毒的基因组以线性的双链DNA形式存在,由蛋白VII和一种称为mu的小蛋白紧密地环绕在其周边,起到类组蛋白样的功效。另一种蛋白V将这些DNA-蛋白复合物连接起来,并借助蛋白VI与病毒衣壳连接在一起。在两条链的5’端各以共价键结合着一个被称为DNA末端蛋白(pTP)复合物(DNA-TPC)的特化的结构,与腺病毒复制密切相关。腺病毒基因组的两端各有一段100bp的逆向末端重复序列(ITR),是复制的起始位点。在左端ITR的3’侧有一段长约300bp的包装信号(ψ)介导腺病毒基因组包装入细菌衣壳。对腺病毒而言,只有包含两端的ITR和包装信号(ψ)的约0.5kb的序列是顺式作用元件,也就是说应该由腺病毒载体自身携带,而其它的30余种蛋白都可以借助辅助病毒(或细胞)反式补足。
腺病毒载体的优点
1.宿主范围广,对人致病性低
腺病毒载体平台可广泛用于人类及非人类蛋白的表达。腺病毒可传染一系列哺乳动物细胞,因此在大多哺乳动物细胞和组织中均可用来表达重组蛋白。特别必须强调的是:腺病毒带有嗜上皮细胞性,而人类的大多数的癌症就是上皮细胞来源的。另外,腺病毒的复制遗传和致敏基因均已非常知道,在人群中已经流行(70-80%成身体内都有腺病毒的中和抗原存在)。人类传染野生型腺病毒后仅造成轻度的自限性病症,且细菌唑治疗有效。
2.在凋亡和非增殖细胞中感染和表达基因
逆转录病毒只能感染增殖性细胞,因此DNA扩增不能在非增殖细胞中进行,而需要使细胞进入大幅培养状况。腺病毒则能传染几乎所有的细胞种类,除了一些抗腺病毒传染的腮腺瘤细胞。腺病毒是研究原代非增殖细胞基因表达的最佳系统,它可以使转换细胞和原代细胞中受到的结果直接进行对比。
3.能有效进行增殖,滴度高
腺病毒系统可生产1010到1011VP/ml的病毒,滴度较高,适用于基因治疗。
4.与人类基因同源
腺病毒载体系统一般应用人类病毒成为载体,以人类细胞成为宿主,因此为人类蛋白进行精确的翻译后加工和适度的折叠提供了一个理想的环境。大多数人类蛋白都可超过高水准表达以及带有完全的功能。
5.不整合到染色体中,无插入致突变性
逆转录病毒可随机整合到寄主染色体,导致遗传失活或激活癌基因。而腺病毒则不仅卵细胞以外几乎在所有已知细胞中都不整合到染色体中,因此不会干扰其他的寄主基因。在卵细胞中融合单拷贝病毒则是造成带有特定特性的转基因动物的一个很好的系统。
6.能在悬浮培养液中扩增
293细胞可以适应悬浮培养,这一调整可使病毒长期扩增。大量事实证明悬浮293细胞可在1~20L的生物反应器中表达重组蛋白。
7.能同时表达多个基因
这是第一个可以在同一细胞株或组织中用来设计表达多个基因的表达系统。最简洁的方式是将带有两个基因的双表达盒插入腺病毒转移载体中,或者用不同的重组细菌共转染目的细胞株来分别表达一个蛋白。测定不同重组病毒的MOI比值可恰当估计各重组蛋白的相对共表达状况。
正是因为具备以上一些特点,腺病毒被如此广泛地应用于体外基因转导、体内接种疫苗、和基因诊断等各个领域。
分类及致病性
自上个世纪50年代发现并成功分离腺病毒以来,已正式看到了100余个血清型,其中人腺病毒有47种,分为A、B、C、D、E和F六个亚群()。基因治疗常见的人的2型及5型腺病毒在抗原学分类上均属C亚群,在DNA序列上有95%的同源性。二者的增殖能力相当强,滴度通常可以超过109PFU/ml,其在单个细胞中的基因组拷贝数可达104(约占细胞总DNA的10%)。病毒颗粒非常稳固,通过CsCl梯度离心可以超过1010~1011PFU/ml,满足动物试验的要求。
2型和5型腺病毒的致病性主要体现为可以避免小孩上呼吸道感染。在感染的最初几天,由于病毒长期复制和积聚,会发生轻度头痛、浑身浮肿、乏力、咽痛等病症。但这种征兆通常是非常短暂而轻微的,在之后的几天里,会随着中和抗体的形成而渐渐消散。虽然可以说高滴度和高免疫原性是腺病毒的主要特征,但也有相对的,如8型腺病毒(主要传染小肠和尿道组织)可以在泄殖腔等淋巴组织中潜伏下来。在C亚群的其它一些腺病毒中也可以看到相同的现象,这是由于病毒的E3区编码的功用可使其躲避宿主的免疫打击。这或许有助于理解何以腺病毒载体在造血系细胞中的基因转导效率显著高于其它组织、细胞。
分类及致病性
腺病毒的生活周期可以分为两个截然不同却又不能割裂开来的两个阶段。第一阶段包括腺病毒颗粒粘附和进入寄主细胞,将基因组释放到宿主细胞核中,以及有选取性地转录和翻译早期基因。在这个阶段,细胞为细菌遗传组复制和腺病毒晚期基因表达并最后传递成熟的感染颗粒腺病毒包装技术,即第二阶段,作好了打算。第一阶段将在6~8个小时内完成,第二阶段则更快,只需4~6个小时。
粘附和进入细胞
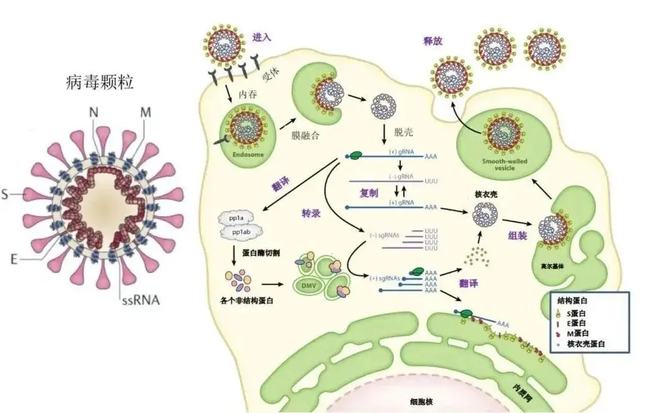
腺病毒传染细胞的过程是从腺病毒纤毛的头节区粘附到细胞表层的特异性受体开始的。因为人腺病毒主要与柯萨奇B病毒共用一种受体,因此这些受体被称为柯萨奇/腺病毒受体即CAR(/)。接出来病毒纤毛基上面五邻体表层的三肽RGD与细胞表层的αvβ3和αvβ5整合素结合,通过内吞作用将腺病毒内化到细胞中并开启溶酶体。在溶酶体的碱性环境下,腺病毒衣壳的氢键将出现变化,被从溶酶体中积聚起来,躲过溶媒体的消化作用。最后,腺病毒颗粒转位到细胞核,通过核孔将细菌DNA释放到细胞核内。相对于脂类体小鼠,腺病毒遗传组进入细胞核是一个特别高效的过程,一般可以超过40%,前者似乎处于胞质的强度与前者比较,而DNA处于细胞核的强度却唯有前者的1/1000。
转录与复制
如果细菌基因组进入细胞核,就将进行一系列的复杂而有序的逐级放大的剪切和转录过程。一般的,以病毒DNA开始复制为分界线,按转录时间的先后,将腺病毒基因大致区别为初期(E1~4)和早期转录单位(L1~5)。各种腺病毒基因又可以进一步地分为更小的转录单位,如E1区可以进一步分为E1A和E1B,每个转录单位都大约有一个另类的开启子。腺病毒基因组进入细胞核后,细胞转录因子首先与E1A区上游的提升子结合,表达E1A蛋白,该多肽的功效是调节细胞代谢,使细菌DNA更容易在细胞中复制。E1A蛋白还可以激活其他初期基因(E1B、E2A、E2B、E3和E4)的推进子,其中E2B驱动另外三个与病毒复制有关的初期基因转录单位末端蛋白前体(,pTP)、单链DNA结合蛋白(-DNA,ssDBP)或者DNA聚合酶的表达,这三个基因的表达产物紧密地结合成一个复合物,与至少三种细胞蛋白相互作用,启动病毒基因组的复制。
一般而言,DNA的复制是由RNA启动的,而在腺病毒却是所谓的蛋白启动(-)。如前所述,腺病毒双链DNA的每条单链的5’端有pTP蛋白结合,pTP通过其Ser-OH与DNA5′端的dCMP5’磷酸之间产生乙酸二脂键。腺病毒的DNA复制首先是以5′端结合有pTP的dCMP作为引物,以3’端的末端反向重复序列(ITR)为模板,进行链置换()合成,置换出的单链分子可以自我退火环化,形成锅柄样环形分子,然后这些环形分子再以同样的体系合成出子代双链DNA分子。
病毒基因组复制一般在感染后数小时开始,同时初期基因的转录和翻译被关闭,晚期基因起初表达。大个别的早期基因的转录是以一个共同的主要早期启动子(MLP,MajorLate)调控的。实际上,MLP的活性与病毒遗传组复制密切相关,有探究表明如果腺病毒遗传组开始复制MLP的活性将显著提高,对此我们将另外行文阐述。晚期基因主要编码腺病毒的结构蛋白。病毒结构蛋白在细胞核内聚集产生真菌衣壳,病毒的基因组被包装出来,形成有传染能力的细菌颗粒,并最后裂解宿主细胞被积聚出去,完成腺病毒的生活周期。腺病毒有显著的种属特异性,人的野生型5型腺病毒(Ad5)传染其他的非人类细胞(如鼠类细胞)后可以表达早期基因,基因组也可有一定程度的复制并无法产生一些不成熟的病毒颗粒,却不能产生成熟的病毒颗粒,也不能二次感染其它细胞。